
编者按:
分类学包括三个关键步骤,分别为特征描述、分类、命名,并且每一步都依赖于前一步。
原核生物的特征描述和分类原则在过去的 50 年中产生了很多变化,直到近些年才形成完整的鉴定体系,现在主要采用多相分类法表型特征和基因型特征相结合的方式,对原核生物进行特征描述和分类。
分子生物学技术的发展使分类鉴定的结果越来越可靠,特别是高通量测序技术的出现,对基因组信息有了更全面、深入和细致的研究。
乳杆菌在食品行业和发酵饲料工业中有着悠久的应用历史。随着科学技术的迅猛发展,人们也逐渐认识到乳杆菌在发酵、生物、农业、医药等多个领域具有重要的应用价值。
早期的乳杆菌分类研究主要依赖于表型特征,如碳源代谢实验、温度、pH、菌落等生长特性对乳酸菌进行分类鉴定。
伴随着分子生物学的诞生和发展,涌现了很多基于 DNA 指纹图谱和核酸序列之外的分类技术。在乳杆菌基因组中,水平基因转移现象是常态,仅以单个或几个基因为研究对象得到的进化关系并不能真实地反映乳杆菌的分类地位和进化关系。
基于全基因组 DNA 序列的基因组学为乳杆菌进化和分类研究注入了强劲动力。全面解析乳杆菌的分类地位、遗传背景和进化历程,对于其开发利用具有极其重要的意义。
刊登于 Microbiome Times 的一篇文章从分类学开始入手,重点关注了乳杆菌属分类的演进,并阐述了重新分类对我们更好地利用乳杆菌来开发产品的重要产业意义。
我们强烈推荐此文,并特别翻译成中文,方便关注益生菌研发和应用的你阅读。
The Lactobacillus Taxonomy change is coming: why, and how to make the most of it?
作者 | Marco Pane and Nina Vinot
编译:Richard
校审:Susan
分类学是研究生物分类的一门科学。达尔文让我们以生命之树的形式来理解物种之间的关系,然而,物种分类本质上是人类精心归纳出来的,反映出的更多是人类思维的结果,并非是物种的真实关系。
Stephen Jay Gould 的一句名言清楚地阐释了分类学与人类思维的关系:我们对生物的分类排序方式反映了我们的思维方式,分类学的历史变迁是我们思维概念转变的直接体现(Gould SJ. 1983)。
因此,对乳杆菌进行预期中的重新分类除了技术和法律上的困难外,还可能会在学术和产业领域引发一场概念革命。
达尔文的《物种起源》中写到的生命之树在动物和植物界已被证明。但是对原核生物(无细胞核的单细胞生物,如细菌和古菌)来说,我们面对的是小到肉眼不可见的生物,它们就需要一种不同的分类方法了。
因为细菌不仅仅会通过分裂将遗传物质传递给后代(基因的垂直遗传),也会在其生活环境中通过不断地与别的细菌融合的方式交换遗传物质(基因的水平转移),尤其是人类肠道中的微生物更是这样(Jeong H,et al.,2019)。它们的生命之树错综复杂,达尔文的进化模型并不适用于这种有大量基因水平转移(HGT)的物种,这种现象对 “生命之树” 的概念建立构成了威胁。
正如 1983 年 Sorin Sonea 在极具前瞻性的《全球的生命体:对细菌的新看法》中所述:“细菌通过获取或交换各种各样的可变遗传系统来发挥功能。” 虽然原核生物出于分类和可视化的目的也采用了 “进化树” 这种简洁明了的结构,但现代观点认为,它们应该抽象成一种复杂的 “生命网”, 而不是“进化树”的形式。
事实上,任何从分类学上对微生物进行分类的尝试都有赖于将特定微生物从生态系统中分离出来,在实验室进行“及时冻结”,这就人为地规避了微生物群落中自然发生的 HGT 现象。换言之,人类观察微生物的活动时,不可避免地限制了其自然的基因组流动性,并就此界定了我们所说的细菌世界中一个物种乃至一个属的概念。
显然,采用新标准将能够揭示以前未见过的生物关系,突出了原核生物分类的动态特征。需要注意的是,分类模型的可靠性与我们解码生物关系的真相所采用的方法是有关的。只有随着分子技术的出现,科学家才有可能绘制细菌的“系统发育树”。
人为对生物进行分类的结果尽管具有一定的随机性,但是对我们来说还是必不可少的,因为分类的核心目的是能对一个客体(抽象或具象)给出一个可以稳定使用的通用名称。
自从 1676 年列文虎克首次用自制显微镜观察到这些 “微小生物” 后,给细菌命名一直是个挑战。它们个体微小,纯培养时不能生长,99.5%的微生物是非培养型的(Harwani D. 2012; Lloyd KG, et al., 2018)。
细菌王国中存在的无性繁殖以及水平基因转移这些难以捉摸的特点让分类学家在细菌定义和命名时纷纷受挫,更恰当地说,细菌王国表现为一种“生命网”(图1),而不是由具有固定显著特征的明确成员组成(Swithers KS, et al., 2009)。
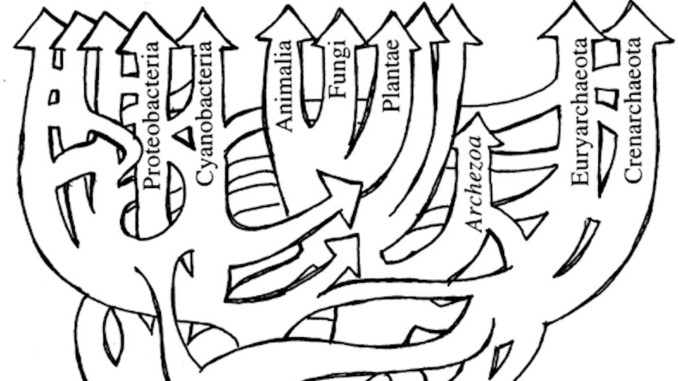
图1:水平基因转移对 “生命之树”的影响,从而形成了一个“生命之网”。(图片来源:Doolittle, 1999)
19 世纪末,巴斯德证实了几种动物传染病的细菌来源,在这个过程中,发现、分离、鉴定致病细菌的技术得以发展。这些致病细菌被称为 pathogens(病原体),这个词由 pathos(疾病)和 gen(成因)组成。在当时,细菌被用作一种治疗手段(疫苗),大部分的研究致力于细菌感染的预防或治疗,以及诊断和免疫。
获益于对致命性传染病的预防和治疗,人类预期寿命得以延长,人口数量开始爆炸性增长。有趣的是,在对抗细菌感染性疾病上获得了成功后,科学家对细菌本身的研究兴趣却在持续下降(而得益于微生物组的研究,才使这一趋势得以恢复)。
这项对病原体的研究让人们掌握了发现、分离、鉴定细菌的技术,这对分离出病原体并且用于治疗疾病至关重要。然而,这也使得临床和实验室中形成了定论:细菌在自然界中是不会发生改变的。
从 19 世纪中叶直到 20 世纪 70 年代,细菌的鉴定与分类绝大多数是依靠表型、形态和化学分类学特征。
20 世纪 50 年代,Carl Woese 对枯草芽孢杆菌(Bacillus subtilis)的孢子进行电离辐射轰击,更深入地观察到了萌发孢子的核糖体和 RNA 结构特征,并与 George Fox 一道提出了“生命三域学说”——古菌域、细菌域、真核生物域(Woese C and Fox G, 1977)。
令人钦佩的是,Woese 在 16S 基因测序技术获得长足发展之前就已使用 rRNA 作为分子标记近 10 年之久,因此他被誉为“16S rRNA 系统进化树之父” (Zhulin IB, 2016)。
随着时间的推移,进化学家提出了不同的进化树模型:最常见的是基于单基因水平的(例如 16S rRNA,编码核糖体小亚基的基因由于在细菌中普遍存在和高度保守,是用来进行分类的最常见基因);最近更多的是基于基因组水平的(由于技术进步,研究者可获得全基因组序列)。
如果没有基于 16S rRNA 这样的基因序列比较获得系统进化模型,我们无法想象目前的细菌分类学会是什么样。
不过,使用 16S rRNA 这样的单基因或许无法反映整个基因组的进化历史。事实上,16S rRNA 大约只占整个基因组的 0.07%,其基因序列中仅有 1.3%的差异与 5000 万年的进化趋异相关(Ochman H,et al., 1999)。使用不同的标记基因会构建出不同的亲缘关系,因此从 16S rRNA 等系统发育标记推算出来的亲缘关系有助于分类,但是却不能代表完整的进化关系。
随着测序和分析技术的快速发展,微生物分类或许开启了第三阶段——系统基因组学时代,这一阶段有赖于全基因组比较(Kislyuk AO, et al., 2011)和蛋白组比较等技术(Callister SJ, et al.,2008)。
此外,还有非系统发育的分类方法。例如表型鉴定,即基于功能相似性进行微生物的分类,而不用考虑其共同祖先。
需要指出的是,尽管基于系统发育的分类学和表型组学都可以用来研究细菌亲缘关系,但它们所解决的问题是不同的。系统发育分类学的任务是重现生命进化的历史进程,而表型组学的任务是根据观察将特征一致的物种归为一类。亲缘关系近的物种通常表型特征相同,但也并不总是这样的(Zhu C, et al., 2015; Sterner B, et al., 2018)。
纵观历史,随着技术和方法的持续进步,科学家们发明了更现代化的工具来厘清原核生物的亲缘关系,这不可避免地给细菌分类学带来了无数次的重新定义。例如 1966 年科学家们进行了一次细菌普查,统计出 28900 种不同物种;仅仅过了 14 年后,由于方法和标准的改进,使得细菌种类缩减到了 1792 种(Skerman VBD, et al., 1980)。到了 2017 年,许多新微生物的物种被鉴定出来,在原核生物标准命名表中(LPSN, www.bacterio.net)的这一数字又上升到了 15626 种。
目前细菌分类学是依据《国际原核生物命名规则》进行的,这个《规则》中涵盖了 65 条细则(Parker CT, et al., 2015),并得到国际认可。

尽管乳杆菌的分类方式发生过变化,但最多也就是在科学范畴内,仅限于是分类学领域中讨论的问题,或者是与病原体鉴定有关的安全问题。
现在,也将是首次,即将到来的乳杆菌分类学变化会对益生菌市场产生前所未有的影响 —— 这是一个快速增长的市场,全世界目前已有数百万人消费益生菌,预计到 2023 年,益生菌的市场规模将达到近 700 亿美元。
目前乳杆菌属已包含 200 多种的菌种,并且还在不断增加,乳杆菌广泛应用于针对健康人群和不健康人群的食品及补充剂中。重新分类可能产生的影响将涉及到安全性、有效性、与科学论文的关系、知识产权,当然还有商业利益。
尽管可能存在对现有市场的重大影响,但是这些菌种之间存在巨大的基因组多样性,这种现状使得分类学家不得不重新审视目前乳杆菌属的分类(图2)。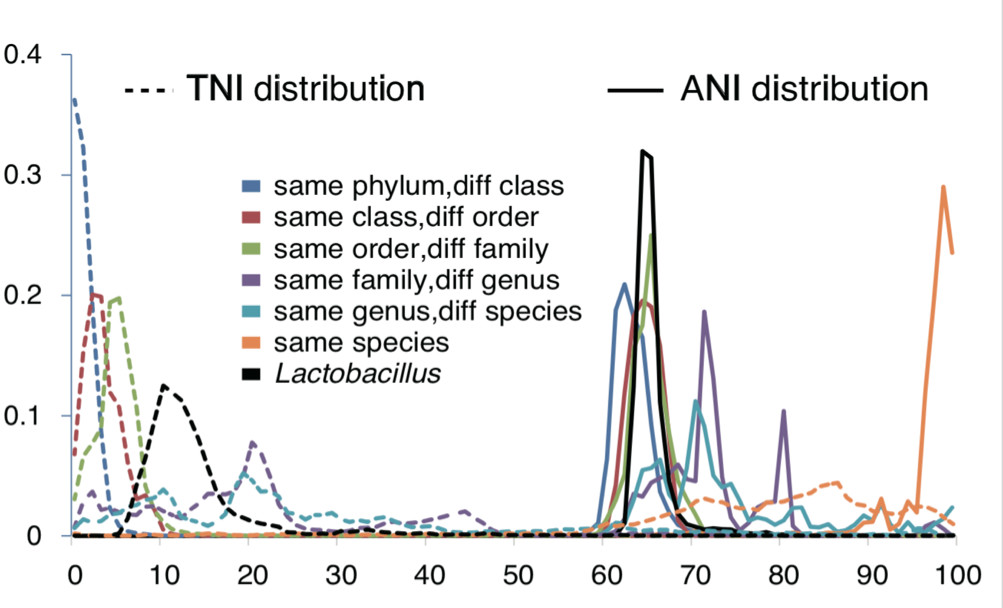 图2:与传统定义的分类单位相比,乳杆菌的 ANI 和 TNI 值的频率分布。
图2:与传统定义的分类单位相比,乳杆菌的 ANI 和 TNI 值的频率分布。
在 TNI 指标中,同一科属的微生物通常具有 20%的遗传相似性,而乳杆菌属的微生物其遗传相似性较低,约为 10%。在 ANI 指标中,同一科属的成员拥有约 70-80%的核苷酸同源性,而目前乳杆菌属的微生物仅有 65%的同源性。
因此分类学家们提出如此建议:应尽快将目前乳杆菌属的 200 多种微生物进行重新分类,归于不同属,并尊重同一科中细菌的相似程度,共同归类于乳杆菌科。
(资料来源:Supplements of Sun Z. et al., 2015, Nature Communications)
而且这种广泛的遗传多样性意味着我们之前鉴定为同属的细菌之间的差别就像人与狐猴的差别一样大(图3)。
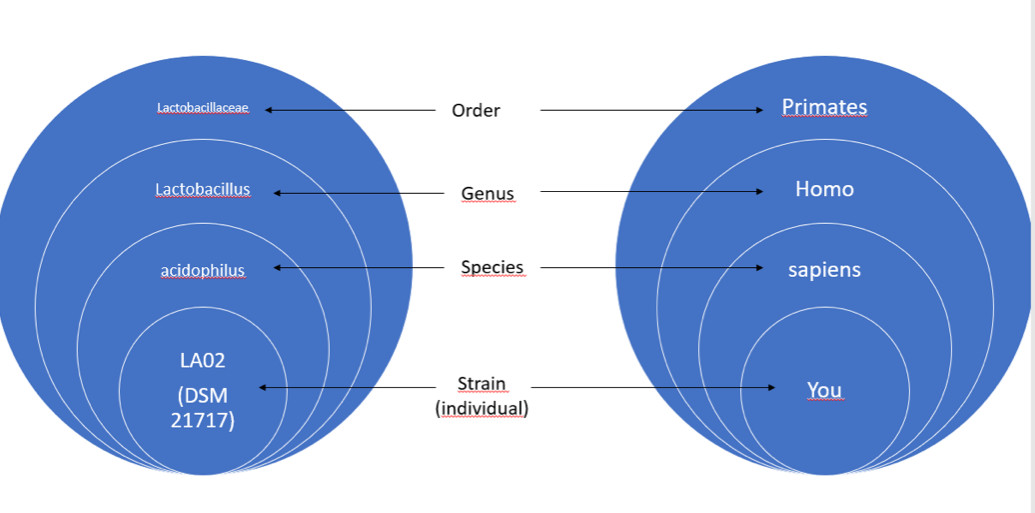 图3:乳杆菌和人科分类的比较
图3:乳杆菌和人科分类的比较

2018年,Salvetti 等人使用距离度量法、ANI、AAI、保守蛋白百分比等不同的方法,对 222 株不同乳杆菌的蛋白质和系统发育标记进行系统性及网络化分析后,强烈建议将乳杆菌属分为 10 个一致性较好的不同菌属,分别为:
(i)德氏乳杆菌(Lactobacillus.delbrueckii),包含嗜淀粉乳杆菌(L. amylophilus)、淀粉样乳杆菌(L. amylotrophicus)和佛罗里达乳杆菌(L. floricola);
(ii)消化乳杆菌(L. alimentarius);
(iii)恶味乳杆菌(L. perolens);
(iv)干酪乳杆菌(L. casei);
(v)清酒乳杆菌(L. sakei)(不包括L. selangorensis);
(vi)棒状乳杆菌(L. coryniformis);
(vii)唾液乳杆菌(L. salivarius);
(viii)植物乳杆菌(L. plantarum);
(ix)罗伊氏乳杆菌(L. reuteri),也包含了牛痘乳杆菌相关的成员(L. vaccinostercus);
(x)布氏乳杆菌(L. buchneri),其中也包括短乳杆菌(L. brevis)、食果糖乳杆菌(L. fructivorans)、丘状乳杆菌(L. collinoides)等成员。
同年,Parks DH 等人在对 94759 个原核微生物基因组的 120 种蛋白质进行系统分析后,提议将乳杆菌属分为 16 个亚群。
之前在 2015 年时,Zheng J 等人在 174 株乳杆菌和小球菌模式株中,基于 172 个涉及生态学和生物化学性质的单拷贝核心基因,得到了 24 个亚群。
最后要说的是,绝对的分类标准是不存在的。使用不同的方法学和判断阈值会得到不同的分类结果,例如在乳杆菌属重新分类的建议上,Salvetti 建议分为 10 个不同属,Parks 建议分为 16 个属,而 Zheng 却分为 24 个属。
分类标准的选择有两个关键点,首先是新属具有更好的同质性,同一属的菌株在相似性上要符合定义标准;最重要的是属的分类数量要稳定,将来发现的新菌种能归到现有属而无需再重新分类。
而且,这些未来更细致的分类单元将有助于我们基于其生理特性、生物地理学、生态学、自然演化史以及与宿主间的潜在互作等方面,研究不同乳杆菌属的共有功能特点,以填补分子作用机制和生态适应性的空白,这有助于我们更好地了解细菌及其用途。
乳杆菌重新分类障碍重重,从标签到质控、技术文件、认证、出口许可、协议、知识产权、再到营销手段、网站等等都需要升级,这些对产业投资者来说都是巨大的投入。
除了投入方面,重新分类给产业带来的另一重大挑战是——尽管用于产品的细菌本身并没有改变,但它们的名称却会改变,这可能会造成与客户及消费者沟通的困难。
但这也将是一个不可多得的机遇,让我们能更清晰地揭示细菌世界,更好地理解新属的特性,开发出功能更有针对性、效果更显著的产品。
细菌的进化史决定了它们如何与宿主互作,这一点也关系到治疗应用中乳杆菌的选择。
例如,植物乳杆菌、干酪乳杆菌、发酵乳杆菌有跟随宿主过游牧的自由生活的历史,这意味着游牧细菌具有更广泛的基因组和更灵活的调节机制,这使其在多变的环境中能生存下来;而罗伊氏乳杆菌、约翰逊乳杆菌、嗜酸乳杆菌是宿主特异性细菌(Duar RM, et al., 2017),与那些未和宿主协同进化的细菌相比,宿主特异性细菌更有竞争力。
如果选择益生菌来对抗病原体,以上这些益生菌非常具有优势。
相反,如果目的是为了刺激免疫系统,那么选择那些未与宿主协同进化的益生菌或许是一个更好的方法,因为这些菌耐受性差,更倾向于刺激免疫反应。
对宿主的适应通常意味着在共生生活方式中一些基因不再被需要,也就渐渐脱落了。有趣的是,在乳杆菌中,宿主特异程度最高的是存在于人类阴道中的乳杆菌,包括惰性乳杆菌(L. iners)、卷曲乳杆菌(L. crispatus)、詹氏乳杆菌(L. jensenii)、加氏乳杆菌(L. gasseri),而惰性乳杆菌和詹氏乳杆菌只在人体阴道中被发现。
这种共生位置特征意味着通过口服可能会影响这些细菌的生存,比如,这些菌的胃酸耐受性和肠道定植性很差,口服会影响其功能的发挥。在欧洲,医疗法案对益生菌产品的监管缺陷引起了人们的质疑,因此 2020 年 5 月前,阴道用益生菌产品都无法获得商业化批准。
通过上面这些例子可以看出,采用新的更细致的分类名称,将有助于在概念上界定微生物的各种特性,从而对有待验证的菌株功能作用提出一个更容易的实验假设。未来更大程度上,透彻地了解这些细菌将有助于我们制定相应的饮食干预策略,使得它们在体内更容易生存,从而真正发挥其特定功能。
根据最新的分类学文献数据,可以推测出乳杆菌属至少能分成 20 个亚群。如果你感兴趣,可以通过这个网站(bacterio.net)一探究竟!
新的分类方法将更加精确地描述具有共同特征的细菌聚类图谱,这是分类学家之间达成共识的结果,而这一结果本身也是技术进步的必然趋势。
重新分类的目标是要确保我们可以用通用且稳定的语言来准确识别细菌。分类学首次对产业产生重大影响,即将到来的乳杆菌重新分类,将影响全球已超过 500 亿美元的益生菌市场,这将是分类学史上重要的历史性时刻。
这也将打开一个新视角,引领我们用新的方法开发益生菌产品。顺便说一下,接下来,双歧杆菌也在等待它们的新面貌!
参考文献:
(滑动文字查看)
Callister SJ, McCue LA, Turse JE, Monroe ME, Auberry KJ, Smith RD et al. (2008). Comparative bacterial proteomics: analysis of the core genome concept. PLoS One 3: e1542.
Ciufo S, Kannan S, Sharma S, Badretdin A, Clark K, Turner S, Brover S, Schoch CL, Kimchi A, DiCuccio M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int J Syst Evol Microbiol. 2018 Jul;68(7):2386-2392. doi: 10.1099/ijsem.0.002809. Epub 2018 May 24. PubMed PMID: 29792589.
Doolittle WF. Phylogenetic classification and the universal tree. Science. 1999 Jun 25;284(5423):2124-9. Review. PubMed PMID: 10381871.
Duar RM, Lin XB, Zheng J, Martino ME, Grenier T, Pérez-Muñoz ME, Leulier F, Gänzle M, Walter J. Lifestyles in transition: evolution and natural history of the genus Lactobacillus. FEMS Microbiol Rev. 2017 Aug 1;41(Supp_1):S27-S48. doi:10.1093/femsre/fux030. Review. PubMed PMID: 28673043.
Gould SJ. 1983. Hen’s teeth and horse’s toes. Norton, New York, NY.
Harwani, Dharmesh. (2012). The Great Plate Count Anomaly and the Unculturable Bacteria. International Journal of Scientific Research. 2. 350-351. 10.15373/22778179/SEP2013/122.
ISO 20128:2006 IDF 192:2006 https://www.iso.org/standard/35292.html
ISO 7889:2003 IDF 117:2003 https://www.iso.org/standard/31880.html
ISO 19344:2015 IDF 232:2015 https://www.iso.org/standard/64658.html
Jeong H, Arif B, Caetano-Anollés G, Kim KM, Nasir A. Horizontal gene transfer in human-associated microorganisms inferred by phylogenetic reconstruction and reconciliation. Sci Rep. 2019 Apr 11;9(1):5953. doi: 10.1038/s41598-019-42227-5. PubMed PMID: 30976019; PubMed Central PMCID: PMC6459891.
Kim M, Oh HS, Park SC, Chun J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol. 2014 Feb;64(Pt 2):346-51. doi: 10.1099/ijs.0.059774-0. Erratum in: Int J Syst Evol Microbiol. 2014 May;64(Pt 5):1825. PubMed PMID: 24505072.
Kislyuk AO, Haegeman B, Bergman NH, Weitz JS (2011). Genomic fluidity: an integrative view of gene diversity within microbial populations. BMC Genomics 12: 32.]
Lloyd KG, Steen AD, Ladau J, Yin J, Crosby L. Phylogenetically Novel Uncultured Microbial Cells Dominate Earth Microbiomes. mSystems. 2018 Sep 25;3(5). pii: e00055-18. doi: 10.1128/mSystems.00055-18. eCollection 2018 Sep-Oct. PubMed PMID: 30273414; PubMed Central PMCID: PMC6156271.
Markets and Markets: https://www.marketsandmarkets.com/Market-Reports/probiotic-market-advanced-technologies-and-global -market-69.html).
Ochman H, Elwyn S, Moran NA. Calibrating bacterial evolution. Proc Natl AcadSci U S A. 1999 Oct 26;96(22):12638-43. PubMed PMID: 10535975; PubMed Central PMCID: PMC23026.
Parker CT, Tindall BJ, Garrity GM. 2015. International Code of Nomenclature of Prokaryotes. Int J Syst Evol Microbiol 68:1825–1829. https:// doi.org/10.1099/ijsem.0.000778.
Parks DH, Chuvochina M, Waite DW, Rinke C, Skarshewski A, Chaumeil PA, Hugenholtz P. A standardized bacterial taxonomy based>Salvetti E, Harris HMB, Felis GE, O’Toole PW. Comparative Genomics of the Genus Lactobacillus Reveals Robust Phylogroups That Provide the Basis for Reclassification. Appl Environ Microbiol. 2018 Aug 17;84(17). pii: e00993-18. doi: 10.1128/AEM.00993-18. Print 2018 Sep 1. Erratum in: Appl Environ Microbiol. 2018 Oct 1;84(20):. PubMed PMID: 29915113; PubMed Central PMCID: PMC6102987.
Skerman VBD, McGowan V, Sneath PHA. Approved lists of bacterial names. Int J Syst Evol Microbiol. 1980;30:225–420.
Sorin Sonea and Maurice Panisset, A New Bacteriology, 1983, Published by Jones and Bartlett Learning.
Soucy SM, Huang J, Gogarten JP. Horizontal gene transfer: building the web of life. Nat Rev Genet. 2015 Aug;16(8):472-82. doi: 10.1038/nrg3962. Review. PubMed PMID: 26184597.
Sterner B, Lidgard S. Moving Past the Systematics Wars. J Hist Biol. 2018 Mar;51(1):31-67. doi: 10.1007/s10739-017-9471-1. PubMed PMID: 28255641.
Sun Z, Harris HM, McCann A, Guo C, Argimón S, Zhang W, Yang X, Jeffery IB, Cooney JC, Kagawa TF, Liu W, Song Y, Salvetti E, Wrobel A, Rasinkangas P, Parkhill J, Rea MC, O’Sullivan O, Ritari J, Douillard FP, Paul Ross R, Yang R,Briner AE, Felis GE, de Vos WM, Barrangou R, Klaenhammer TR, Caufield PW, Cui Y, Zhang H, O’Toole PW. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat Commun. 2015 Sep 29;6:8322. doi: 10.1038/ncomms9322. PubMed PMID: 26415554; PubMedCentral PMCID: PMC4667430.]
Swithers KS, Gogarten JP, Fournier GP. Trees in the web of life. J Biol. 2009;8(6):54. doi: 10.1186/jbiol160. Epub 2009 Jul 13. Review. PubMed PMID: 19664165; PubMed Central PMCID: PMC2737374.
Woese CR, Fox GE.1977. Phylogenetic structure of the prokaryoticdomain: the primary kingdoms. Proc Natl Acad SciUSA74:5088 –5090.http://dx.doi.org/10.1073/pnas.74.11.5088.
Zheng J, Ruan L, Sun M, Gänzle M. A Genomic View of Lactobacilli and Pediococci Demonstrates that Phylogeny Matches Ecology and Physiology. Appl Environ Microbiol. 2015 Oct;81(20):7233-43. doi: 10.1128/AEM.02116-15. Epub 2015 Aug 7. PubMed PMID: 26253671; PubMed Central PMCID: PMC4579461.
Zhu C, Delmont TO, Vogel TM, Bromberg Y. Functional Basis of Microorganism Classification. PLoS Comput Biol. 2015 Aug 28;11(8):e1004472. doi: 10.1371/journal.pcbi.1004472. eCollection 2015 Aug. PubMed PMID: 26317871; PubMed Central PMCID: PMC4552647.
Zhulin IB. 2016. Classic spotlight: 16S rRNA redefines microbiology.J Bacteriol 198:2764-2765. doi:10.1128/JB.00616-16
本文来源:http://www.microbiometimes.com






